Article Text

Abstract
As of 25 January 2022, over 349 million individuals have received a confirmed diagnosis of covid-19, with over 5.59 million confirmed deaths associated with the SARS-CoV-2 virus. The covid-19 pandemic has prompted an extensive global effort to study the molecular evolution of the virus and develop vaccines to prevent its spread. Although rigorous determination of SARS-CoV-2 infectivity remains elusive, owing to the continuous evolution of the virus, steps have been made to understand its genome, structure, and emerging genetic mutations. The SARS-CoV-2 genome is composed of several open reading frames and structural proteins, including the spike protein, which is essential for entry into host cells. As of 25 January 2022, the World Health Organization has reported five variants of concern, two variants of interest, and three variants under monitoring. Additional sublineages have since been identified, and are being monitored. The mutations harboured in these variants confer an increased transmissibility, severity of disease, and escape from neutralising antibodies compared with the primary strain. The current vaccine strategy, including booster doses, provides protection from severe disease. As of 24 January 2022, 33 vaccines have been approved for use in 197 countries. In this review, we discuss the genetics, structure, and transmission methods of SARS-CoV-2 and its variants, highlighting how mutations provide enhanced abilities to spread and inflict disease. This review also outlines the vaccines currently in use around the world, providing evidence for every vaccine's immunogenicity and effectiveness.
- Covid-19
- COVID-19
- Virology
Data availability statement
Data are available in a public, open access repository.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Introduction
Seven coronaviruses can infect humans, all belonging to the alpha or beta subgroups, including 229E (alpha), NL63 (alpha), OC43 (beta), and HKU1 (beta).1 Over the past two decades, three notable beta coronaviruses (severe acute respiratory syndrome coronavirus (SARS-CoV) in 2002; Middle East respiratory syndrome coronavirus (MERS-CoV) in 2011; and most recently, severe acute respiratory syndrome 2 (SARS-CoV-2) in 2019) have emerged and caused severe illness, resulting in debilitating disease and worldwide deaths. SARS-CoV-2 is the pathogen responsible for the current coronavirus 2019 (covid-19) pandemic and has caused more than 5.59 million deaths in around two years and resulted in multisystem illness in several million people.2
All viruses change and mutate over time, with most changes having little to no impact. However, some mutations could alter its pathogenic or transmission potential and might, therefore, increase disease severity or hinder the effectiveness of vaccines and therapeutic strategies. The World Health Organization3 classifies variants of concern as SARS-CoV-2 variants that increase transmissibility, disease severity, or virulence or that decrease the effectiveness of public health measures, diagnostics, therapeutics, or vaccines. Variants of interest are variants with genetic changes predicted to enhance the virulence and transmissibility of the virus, which have been identified to cause community transmission in multiple countries and pose a possible risk to global public health. Lastly, variants under monitoring are those with genetic changes are suspected to affect virus characteristics and have currently unclear phenotypic or epidemiological effects. Variants under monitoring are not typically assigned a name until they are upgraded to variants of interest or concern. The full working definitions of variants of concern, variants of interest, and variants under monitoring can be found on the WHO website for tracking SARS-CoV-2 variants (www.who.int/en/activities/tracking-SARS-CoV-2-variants/).3 As of 25 January 2022, WHO reports five variants of concern (alpha, beta, gamma, delta, and omicron), two variants of interest (lambda and mu), and three variants under monitoring.3 Former variants of concern, variants of interest, or variants under monitoring have been reclassified as "formerly monitored variants," owing to these variants no longer circulating, having little impact on the epidemiological situation, or having no concerning properties.3 Since the beginning of the covid-19 pandemic, the rapid development of effective covid-19 vaccines has taken place around the world. As of 24 January 2022, 33 vaccines have been approved for use in 197 countries, with 10 vaccines having gained emergency use listing approval from WHO.4
In this review, we provide an overview of the genome and structure of SARS-CoV-2, describing how these elements allow the virus to infect and replicate inside of host cells, before outlining how certain mutations harboured by SARS-CoV-2 variants enhance these abilities. Next, we examine the current state of vaccine development around the world and provide evidence of the effectiveness of booster doses.
Sources and selection criteria
We searched PubMed and Embase databases for covid-19 related articles published between 1 January 2020 and 25 January 2022 and for general coronavirus related articles published from 1 January 2000 onwards. Our search terms included SARS-CoV-2, covid-19, and specific terms including virology, genome, variants, and vaccine. Additional, specific search terms are outlined in online supplemental file 1. We performed further manual searching for additional articles and data using relevant databases, including who.int, gov.uk, and ecdc.europa.eu/en. Owing to the rapidly evolving nature of the literature involving SARS-CoV-2, we also searched preprint databases including MedRxiv and BioRxiv. We selected studies through different criteria (online supplemental file 1), owing to the various topics discussed here. Overall, studies were selected on the basis of quality and impact factor of publishing journal, with real world studies with large sample sizes of the greatest interest.
Supplemental material
Viral transmission, clinical presentation, and genetic susceptibility of covid-19
SARS-CoV-2 is predominantly spread via respiratory droplet transmission, spreading between people through close contact, coughing, or sneezing. The virus can also spread through airborne transmission, fomite transmission, and via other modes, such as through biological material including urine and faeces.5 6 The SARS-CoV-2 virus can survive on surfaces or survive suspended in air droplets for long periods. Indeed, on plastic, stainless steel, and glass surfaces, the half life of the virus is around 5.3, 4.4, and 4.2 hours, respectively,7 with no difference seen between SARS-CoV-2 variants.8 Although SARS-CoV-2 can be detected on inanimate surfaces for hours and days, owing to the evaporation of water droplets (the viruses’ living environment), the concentration of the virus plummets rapidly.9 Protective measures, including use of personal protective equipment, maintenance of indoor ventilation, and disinfection hands and surfaces, can effectively limit the spread of SARS-CoV-2.10
Once inside the airways, SARS-CoV-2 can infect ciliated, mucus secreting, and club cells of bronchial epithelium, type 1 pneumocytes within the lungs, and the conjunctival mucosa.11 The clinical presentation of covid-19 is non-specific and heterogeneous, and infection can result in a wide spectrum of symptoms. After an incubation period of 4-14 days, symptoms range from mild to severe disease and, in some instances, can result in death.12 The most common covid-19 symptoms include fever, cough, dyspnoea, and fatigue,13 14 while myalgia, gastrointestinal issues, cognitive deficits, and other symptoms are reported. Asymptomatic individuals can also test positive for covid-19.15 16 Although the entire population is susceptible to covid-19 infection, some subgroups within the general population are more susceptible to developing poorer clinical outcomes.
Risk factors associated with increased probability of hospital admission, severe disease, and fatal outcome with covid-19 have been identified. Older age17–19; male sex20 21; belonging to an ethnic minority group21 22; and comorbidities including diabetes, hypertension, and lung disease,18 23–25 malignancy, and immunodeficiency26–28 have all been associated with more severe covid-19. The duration and treatment of covid-19 symptoms will also have profound influences on the severity of disease and the acute and long term outcomes after recovery. The host genetic background is thought to have an influence on the susceptibility and severity of covid-19, possibly explaining the broad spectrum of clinical manifestations that can develop in seemingly similar individuals. A meta-analysis, consisting of 49,562 patients with covid-19 across numerous ancestry groups, identified four gene loci associated with susceptibility to covid-19 (SLC6A20, RPL24, ABO, PLEKHA4) and nine associated with increased risk of severe covid-19 (LZTFL1, FOXP4, TMEM65, OAS1, KANSL1, TAC4, DPP9, RAVER1, and IFNAR2).29 Meanwhile, genome wide association studies spanning across Europe, the US, and the UK identified a gene cluster on chromosome three (chr3p21.31) as being strongly linked with susceptibility and severity of covid-19.30 31 Polymorphisms in the genes of the angiotensin converting enzyme 2 (ACE2) receptor and transmembrane protease serine 2 (TMPRSS2) have also been shown to enhance SARS-CoV-2 viral entry,32 33 with differential polymorphisms seen across ethnic minority populations, which might partly explain why certain ethnic groups are more susceptible to severe covid-19. Increased ACE2 receptor levels have also been associated with other risk factors of covid-19, including smoking and increasing age.34 The use of polygenetic risk scores might be useful in determining an individual’s risk for developing severe disease caused by covid-19.35 A polygenetic risk score infers a person’s risk of susceptibility to, or development of, a certain disease based on the total number of genomic variations they possess. Determining polygenetic risk scores with the inclusion of comorbidities, such as chronic obstructive pulmonary disease,36 or other aspects such as coagulation factors,37 could improve the usefulness of these scores in determining a person’s risk of severe covid-19.
Virology of SARS-CoV-2
SARS-CoV-2 is a positive stranded RNA virus belonging to Coronaviridae family. Coronaviruses, which have crown-like appearances, are the largest known RNA viruses and are thought to primarily infect vertebrates.38 39 SARS-CoV-2 belongs to the beta genus of the coronaviruses and has a genome size varying from 29.8 to 29.9 kb.40 Human coronavirus genomes consist of a variable number of open reading frames (ORFs). Following the typical 5’ to 3’ order, the beginning two thirds of the SARS-CoV-2 genome contains two ORFs (ORF1a and ORF1b) that, inside the host cell, are translated at the rough endoplasmic reticulum into polyprotein 1a (pp1a) and polyprotein 1ab (pp1ab), respectively.40 These polyproteins are cleaved into 16 non-structural proteins (nsp): nsp1-11, from pp1a; and nsp12-16, from pp1ab. The proteolytic release of nsp1 occurs rapidly, which enables it to interfere with translation processes of the host cell by inducing cellular mRNA degradation.41–43 Nsp2-16 contain the viruses’ replication and transcription complex and encode multiple enzymes with many functions, including proteases, helicase, polymerase, exonuclease and endonuclease, N7-methyltransferase and 2’O-methyltransferase, and de-ubiquitination enzymes.44 45
The final third of human coronavirus genomes contain genes that encode structural and accessory proteins. The four major structural proteins encoded here are the nucleocapsid (N), membrane (M), envelope (E), and spike glycoprotein (S) proteins.46 47 The N protein is associated with the viral RNA genome, is involved in RNA synthesis regulation, and interacts with the M protein during viral budding.39 48 The M protein is important for viral assembly, it contains a short N-terminal domain that projects onto the external surface of the envelope and a long internal C-terminal.39 The E protein function is largely unknown; however, along with the N and M proteins, it is required for viral assembly and release.47 Lastly, the S protein gives coronaviruses their characteristic spikes that compose their crownlike appearance. This protein projects through the viral envelope, is heavily glycosylated, and regulates host cell membrane receptor binding and fusion of the viral and cellular membrane.49 The functions of the 11 accessory proteins encoded within the one-third closest to the 3’ end of the SARS-CoV-2 genome are not fully understood. These accessory proteins are encoded by the ORF3a, ORF3b, ORF3c, ORF3d, ORF6, ORF7a, ORF7b, ORF8, ORF9b, ORC9c, and ORF10 genes. Some of these proteins, including ORF3b, ORF6, ORF7a, and ORF8, are interferon antagonists that impair the host cell immune response,50–53 whereas ORF3a might promote virus release54 and is involved in apoptosis of host cells through caspase-3 activation.55 ORF9b and ORF9c are known to suppress the host antiviral response by interacting with host cell organelles,56–58 whereas a clear understanding of the functions of ORF3c, ORF7b, and ORF10 remains unclear.59 Figure 1 (A,B) depicts the genome and structure of SARS-CoV-2.

Genome and structure of SARS-CoV-2. (A) SARS-CoV-2 genome and spike (S) protein amino acid composition. The SARS-CoV-2 genome is about 30 000 base pairs (bp) long and consists of open reading frames (ORF) and elements that are essential for the virus’ structure. The S protein is responsible for binding and entry into host cells. SARS-CoV-2 variants of concern contain various S protein non-synonymous mutations that result in amino acid changes in the receptor binding domain (orange bracketed text) and the S1/S2 subunit interface (black bracketed text), which have been shown to enhance transmissibility of the virus. Variants of concern include alpha (α), beta (β), gamma (γ), delta (δ), and omicron (O). (B) SARS-CoV-2 structure. SARS-CoV-2 is an RNA virus that has a crown-like appearance and contains four major structural proteins: nucleocapsid (N), spike (S), envelope (E), and membrane (M). (C) Viral S protein and human angiotensin converting enzyme 2 (ACE2) interaction. The SARS-CoV-2 S protein directly interacts with human ACE2 receptors in order to gain entry into host cells. The receptor binding domain (RBD) of the S protein tightly binds to ACE2. (D) S protein structure. The S protein protrudes out from the main SARS-CoV-2 bulk and is comprised of two subunits: S1 and S2. S1 contains the RBD, which directly interacts with the human ACE2 receptor, while the S1/S2 interface contains a furin cleavage site that is cleaved to allow S2 to fuse with the host cell membrane. Both the RBD and the S1/S2 interface contain transmissibility increasing mutations that are harboured in variants of concern
The S glycoprotein is composed of two functionally distinct subunits (S1 and S2) and is essential for viral entry into host cells. The N-terminal S1 domain of the protein contains the receptor binding domain (RBD) that directly interacts with the ACE2 receptor on the host cell, which is the primary receptor that SARS-Cov-2 uses for cell entry.60 The C-terminal S2 domain fuses the host and viral membranes to allow for entry of the viral genome into the host cell.61 The subunits of the trimeric S complex are either in a closed (pre-fusion stage) or open (post-fusion stage) conformation,62 with one subunit always in an open conformation to allow for ACE2 recognition and binding.63 The RBD itself consists of five anti-parallel β strands surrounded by several α helices.64 From closed to open conformation, the RBD undergoes structural rearrangement whereby the globular head region rotates clockwise, which alters is elecropotential surface.64 Once positioned, numerous residues within the RBD form either hydrogen bonds or salt bridges with residues of the ACE2 receptor, allowing for tight binding,65 while the concave structure of the RBD allows for three distinct binding regions.64 Following binding between the S protein and the host cell receptor, host cell proteases cleave the S protein, causing the release of the S2 domain which allows for fusion and cell entry.66 Figure 1 (C,D) shows the structure and function of the S protein.
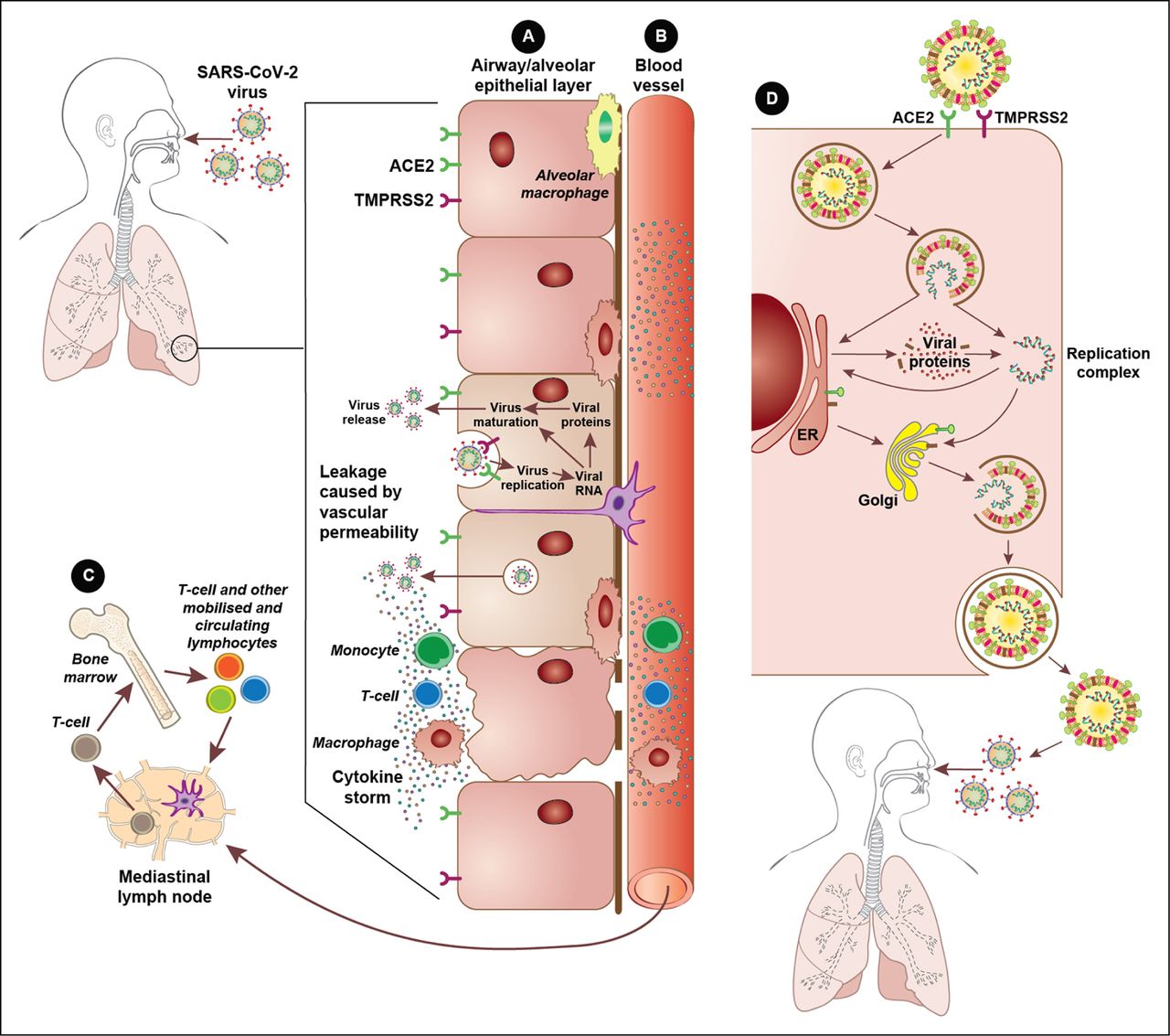
The ACE2 receptor is expressed in numerous cell types throughout the human body, including in the lungs, oral and nasal mucosas, heart, gastrointestinal tract, kidneys, liver, spleen, and brain,67 highlighting the widespread infection that SARS-CoV-2 can inflict. Meanwhile, TMPRSS2, a host cell protease, facilitates fusion of the viral and host cell membranes,68 and could have a role in the spread of the virus in the airways.68 Host cell cathepsin L might also aid in SARS-CoV-2 cell entry by cleaving the S protein.69 Indeed, a clinically approved protease inhibitor has been shown to block SARS-CoV-2 cell entry.70 Figure 2 depicts the mechanism by which SARS-CoV-2 gains entry into and replicates inside host cells, and summarises the host cell immune response.

{kind=link}
{kind=link}
Viral entry and host response. (A) At the alveolar epithelial cell layer. Epithelial cells in the lungs express both angiotensin converting enzyme 2 (ACE2) receptors and transmembrane protease serine 2 (TMPRSS2), allowing for infection by SARS-CoV-2. Replication of the virus within these cells induces an intense immune response that attracts monocytes, T cells, and macrophages and, in some instances, can result in a cytokine storm. (B) Within nearby blood vessels. Cytokines produced by the epithelial cell layer are released into blood vessels supplying the infected tissue, which causes the recruitment of further immune cells to the area, driving the damaging inflammatory response further. Circulating cytokines also create a systemic inflammatory environment. (C) Adaptive immune response. Circulating lymphocytes carry viral antigens to lymph nodes and bone marrow to begin the adaptive immune system processes whereby B cells, and later antibodies, are activated. (D) SARS-CoV2 host replication. The SARS-CoV-2 virus uses the ACE2 receptor and TMPRSS2 to gain entry into human cells. Following release of the viral RNA within the host cell, the virus uses the host endoplasmic reticulum (ER) and Golgi apparatus to produce and manufacture new viral particles, which are released out of the cell to infect other cells and new hosts
Variants of SARS-CoV-2
Most viral mutations have a limited impact on the viruses’ ability to infect, replicate, escape host immunity, and transmit; however, certain mutations can give a viral strain a competitive advantage and, through natural selection, give it the ability to become dominant. Many mutations observed in SARS-CoV-2 variants are found within the RBD or the N-terminal domain of the S protein, which alters the three dimensional structure of the S protein. Not only can these changes affect the transmission abilities of the virus, but it can also allow it to better escape the immune response, such as from neutralising antibodies either elicited through vaccine administration or natural infection.
The SARS-CoV-2 virus has mutated numerous times, with estimates suggesting that circulating lineages acquire nucleotide mutations at rates of around one to two mutations per month.71 The current method of identifying variants relies on the use of genomic testing such as whole genome sequencing, partial S gene sequencing, or assays based on nucleic acid amplification.72 The aspects of different variants that most people experience, however, is the clinical symptoms they inflict. Certain variants (eg, alpha, delta) induce a greater risk of severe disease and death,73 while others (eg, omicron) are more likely to induce milder symptom.74 75 Moreover, individual symptoms can differ between variants. For example, the gamma variant is associated inflicting anosmia and dysgeusia,76 which is less commonly seen in omicron infections. Moving forward, the clinical themes and symptoms associated with emerging variants should be elucidated rapidly so that the public and healthcare professionals can rapidly identify possible cases of covid-19.
WHO has tracked and monitored SARS-CoV-2 variants since the covid-19 pandemic began to identify variants of concern. As of 25 January 2022, WHO reported five variants of concern, two variants of interest, and three variants under monitoring (table 1).3 Here, we report studies that compare SARS-CoV-2 variants to the primary virus strain. The primary strain is the strain of the virus that first emerged in Wuhan, China at the end of 2019 and spread around the world in the first wave of infections, which is often also referred to as the Wuhan-Hu-1, B.1, or wildtype strain.
SARS-CoV-2 variants and related spike protein mutations
Variants of concern
Alpha variant B.1.1.7
The alpha SARS-CoV-2 variant of the B.1.1.7 lineage was first documented in the UK in September 2020 and classified as a variant of concern on 18 December 2020.3 77 This variant contains S protein mutations that have potential biological effects. Firstly, the S protein residue 501, a key contact residue within the RBD, forms a portion of the binding loop in the contact region of the ACE2 receptor, forms a hydrogen bond with the Y41 residue of the ACE2 receptor, and stabilises the ACE2 K353 residue.65 78 79 The alpha variant has an N501Y mutation, which increases the binding affinity of the RBD to the ACE2 receptor.80 Next, the P681H mutation contained within the alpha variant is located immediately adjacent to the 682-685 furin cleavage site, at the interface of the S1 and S2 domains.81 The S1/S2 furin cleavage site prompts entry into respiratory epithelial cells and partly determines the transmissibility of the virus,82–84 while the P681H mutation makes the furin cleavage site less acidic, meaning it is more effectively recognised and cleaved.85 86 Alpha also contains a D614G mutation, located within the S1/S2 furin cleavage site, which increases SARS-CoV-2 binding affinity to the ACE2 receptor and increases infectivity.87 Other mutations within the alpha variant enhance the ability of the virus to escape antibody detection, such as the two amino acid deletion at sites 69-70 in the N-terminal domain of the S protein,88 89 while other mutations show limited or no effects.90 In February 2021, viruses of the B.1.1.7 lineage with the added S protein mutation E484K were identified, which could have threatened vaccine effectiveness owing to the mutation conferring an increased resistance to neutralising vaccine elicited and monoclonal antibodies.91 This mutation had limited effects, however, and variants containing it failed to dominate.
Epidemiological studies have explored the alpha variant, with a case-control study of 27 633 respiratory samples originating from 20 primary care centres in Madrid, Spain, finding that the probability of admission to an intensive care unit was twice as high in patients infected with the alpha variant compared with those infected with the primary strain.92 Furthermore, this variant became the dominant strain within four months, and led to an increase in disease burden as a result.92
Meanwhile in Cannes, France, infection with the alpha variant was associated with a 3.8-fold higher risk of transfer to intensive care or death compared with the primary strain, as determined through a retrospective cohort study of 158 patients with covid-19.93 A large retrospective cohort study including a total of 476 973 participants found that, during the third covid-19 wave in Canada, where 91% of infections were caused by the alpha variant, the risk of both hospital admission (adjusted odds ratio 1.57) and death (1.52) was higher than primary strain infections.94 Overall, the alpha variant was about 50-70% more transmissible and was associated with a 30-60% increased risk of hospital admission and death compared with the primary strain.95–100
The alpha variant was found to have a minimal impact on the effectiveness of current vaccines,101 102 while the risk of reinfection remained similar for this variant as with previous ones.103 On 3 September 2021, the European Centre for Disease Prevention and Control (ECDC) reclassified the alpha, and the alpha +E484K mutation variants from a variant of concern to a de-escalated variant.104
Beta variant B.1.351
The beta SARS-CoV-2 variant, of the B.1.351 lineage, was first documented in South Africa in May 2020.3 This variant contains five S protein mutations of interest: N501Y, E484K, D614G, K417N, and A701V. Like the alpha variant, the beta variant contains the mutations N501Y, E484K, and D614G, which increase ACE2 receptor binding affinity,80 87 increase virulence,105 and enhance resistance to neutralising antibodies.91 106 The K417 residue of the SARS-CoV-2 S protein interacts with the D30 residue of the ACE2 receptor, forming a salt bridge across the central contact region,65 78 although the K417N mutation appears to have a limited impact on ACE2 receptor binding.80 The A701V mutation is located close to the furin cleavage site but has a minimal impact on transmissibility or antibody resistance.101
In a genomic and epidemiological study, researchers concluded that the beta SARS-CoV-2 variant had a selective advantage over previous variants from its increased transmissibility and immune escape abilities,107 108 whereas the E484K/N501K mutations enhanced the binding affinity of the beta variant and, hence, increased its transmissibility.109 A retrospective cohort study of 22 068 participants found that infection with the beta variant was associated with an increased risk of hospital admission compared with an infection with a non-variant of concern (hazard ratio 2.30).100 Overall, the beta variant is about 25-50% more transmissible, is associated with a possible increase in risk of hospital mortality, and has enhanced resistance to antibody neutralisation compared with previous variants.107 108 110
Gamma variant P.1
The gamma variant is of the P.1 lineage and was first reported in November 2020 from travellers returning to Japan from Brazil, and was later discovered in Brazil.3 111 This variant contains the following S protein mutations of interest: K417T, E484K, N501Y, D614G, and H655Y.104 As mentioned, the N501Y and D614G mutations increase both ACE2 receptor binding affinity and infectivity of the virus.80 87 The N501Y, K417N/T, and E484K mutation trinity, meanwhile, is shared by both gamma and beta variants, and is associated with enhanced infectivity and lethality compared with the N501Y mutation alone, possibly from tighter binding of the S protein to the ACE2 receptor due to increased electrostatic contribution.112 The gamma variant also includes the H655Y mutation, which was found to provide enhanced viral escape abilities from multiple human monoclonal antibodies in vitro.113
The gamma variant is associated with heightened transmissibility,109 110 114 with one study concluding that it possesses a 1.7-fold to 2.4-fold increased transmissibility compared with previous variants.115 Additionally, the wave of infections caused by the gamma variant in Brazil was associated with a 13% increase in death rate compared with the previous wave, suggesting the greater virulence held by the gamma variant than by previous viral strains.116
A surveillance study from seven European countries concluded that the gamma variant was associated with a higher risk of admission to hospital (adjusted odds ratio 2.6) and intensive care (2.2) when compared with cases of non-variants of concern.117 In Manaus, Brazil, the resurgence of covid-19, despite high seroprevalence, suggested that the gamma variant had a moderate resistance to neutralising antibodies,118 however, the variant has been shown to be significantly less resistant to neutralising antibodies than other variants, including the beta variant.119
Delta variant B.1.617.2
The delta variant, from the B.1.617.2 lineage, was first documented in India in October 2020 and was classified as a variant of concern on 11 May 2021.3 The S protein mutations of interest P681R and D614G are also located in the delta variant104 and similarly affect its ACE2 receptor binding affinity and transmissibility.106 120 121 Unlike the E484K mutation seen in previous variants, the delta variant contains the E484Q mutation that, along with a L452R mutation also located within the RBD, causes significantly higher affinity for the ACE2 receptor than the primary strain or the E484K mutation alone.122 The L452R mutation alone results in greater RBD-ACE2 receptor binding affinity and enhanced escape from neutralising antibodies.123 124 Lastly, the delta variant contains the T478K mutation, located on the interface between the S protein and the ACE2 receptor when bound, which increases the electrostatic potential of the S protein and enhances binding affinity.125
The delta variant quickly became the dominant variant in the UK,126 US,127 Europe, and around the world.128 The mutations present in the delta variant enhanced the transmissibility of the virus as a result of increased binding affinity to the ACE2 receptor.109 The reproduction number of the delta variant is estimated to be 97% greater than that of non-variants of concern or non-variants of interest, and about three times that of the alpha, beta, and gamma variants.110 This increased reproductivity highlights the delta variant's competitive advantage over earlier ones and how it rapidly became the dominant strain globally. The fast replication rate of delta probably contributes to its increased transmissibility compared with the alpha, beta, and gamma variants. In a cohort study consisting of 167 infections, the delta variant could be detected by polymerase chain reaction within the first four days from exposure, whereas non-delta covid-19 infections could be detected after only six days.129 Furthermore, people infected with the delta variant were found to have significantly higher viral loads than people infected with other strains,129 including the beta variant.130 The delta variant is also thought to better escape neutralisation, with the frequency of post-vaccination infections much higher for the delta variant than infections with the primary strain in India,131 and blood serum samples from individuals who had received one dose of a covid-19 vaccine showing minimal neutralisation of the delta variant.132
The delta variant is also associated with an increased disease severity. In Scotland, infection with the delta variant was associated with an increased risk of hospital admission (hazard ratio 1.85) compared with infection with the alpha variant.133 Compared with infections involving non-variants of concern, North American retrospective cohort studies showed that infection with the delta variant was associated with a 108%134 or hazard ratio of 2.28 (95% confidence interval 1.56-3.34) 100 increased risk of hospital admission, a 234% increased risk for admission to intensive care, and a 132% increased risk of death.134 Lastly, in a cross sectional study of 6238 individuals infected with the delta variant and 3262 infected with the primary strain in India, researchers found that the risk of death was around 1.8 times higher for delta infections, while the delta variant also infected and induced symptoms in a greater proportion of younger people (age 0-19 years) than did the primary strain.131
Omicron variant B.1.1.529
The omicron variant is of the B.1.1.529 lineage and was first discovered in November 2021 in South Africa and Botswana before being detected in multiple countries and classified as a variant of concern on 26 November 2021.3 This variant contains over 30 S protein mutations,104 23 of which have been previously identified, including K417N, T478K, E484A, D614G, H655Y, P681H, and N501Y.135 Fifteen omicron mutations are contained within the RBD,17 providing the variant with a substantially enhanced binding affinity to the ACE2 receptor.135 136 In addition, various single mutations in the RBD of the omicron variant impair the effectiveness of neutralising antibodies, including K417N, N440K, G446S, E484A, Q493K, G496S, G339D, S371L, and S375F.17
The emergence of omicron has been followed by a surge of infections worldwide. Early data from South Africa have indicated that the proportion of covid-19 infections caused by the omicron variant rose from 3% in early October 2021 to 98% by early December 2021.137 In late December 2021, meanwhile, the doubling time for the number of omicron infections was between two and three in the UK, US, and much of Europe,138 139 highlighting the transmissibility of this variant. The mutations in the omicron variant that enhance its binding affinity135 136 and ability to escape neutralising antibodies17 probably drove its rapid spread, as did its fast replication rate, which is around 70 times faster than the delta and primary strains.140 The reinfection rate of the omicron variant has also been found to be more than ten times higher than that of previous variants in studies from Scotland141 and South Africa.142
The omicron variant has extensive but incomplete escape abilities from naturally acquired and vaccine induced immunity.143 144 Compared with the delta variant, the omicron variant needs around a 10-fold increased antibody titre to be neutralised, after vaccination with either the ChAdOx1 nCoV-19 (Oxford-AstraZeneca) or BNT162b2 (Pfizer-BioNTech) vaccines.145 Indeed, blood serum from individuals who had received two doses of the BNT162b2 vaccine showed more than a 25-fold reduction in neutralising antibody titres against the omicron variant compared with the primary strain.146 T cell responses to the omicron variant could remain intact, however. Data from one preprint study indicated that 70-80% of the T cell response targeting the S protein was maintained in those individuals vaccinated or with previous infection, while the magnitude of T cells cross reacting with the omicron variant was similar to that of both delta and beta variants.147 Furthermore, data from Pfizer-BioNtech revealed that 80% of the epitopes in the omicron variant S protein that are recognised by CD8 T cells were not affected by the variant’s mutations, after two doses of the vaccine.146 T cell responses induced from vaccination or prior infection could, therefore, provide some protection from severe disease.
Recent real world evidence has implied that omicron infection is milder in severity than previous variants. In an early South African analysis, the risk of hospital admission (adjusted odds ratio 0.2) was lower for omicron infections than for non-omicron infections,137 while omicron infected individuals had a lower risk of severe disease than delta infected individuals (0.3).137 In December 2021 in England, omicron infections were found to induce a greatly reduced risk of hospital admission or presentation for emergency care than delta infections.74 75 The decreased disease severity inflicted by the omicron variant could be due to its reduced capacity for replication in lung tissue, which was found to be more than 10 times less in lung tissue than the delta variant.140 Concordantly, the S protein of the omicron variant is less efficient at cleaving the ACE2 receptor and entering cells of lung organoids,145 and is also less able to cause fusion between lung cells than the S protein of the delta variant,145 which is often observed in severe covid-19. The reduction in replication within the lungs, and the preservation of T cell responses probably contribute to the milder disease exerted by the omicron variant.
The original Omicron variant is referred to as BA.1, due to the detection of several sublineages of the variant in circulation. While the emergence of BA.1 coincided with a wave of covid-19 infections around the world due to its higher transmissibility and increased risk of reinfection than previous variants,148 sublineages BA.2 and BA.3 are also circulating, with BA.2 now responsible for an increasing number of the reported cases.149 The current data remains limited, however, the UK Health Security Agency report that BA.2 has an increased growth rate compared to BA.1 although this report did not find any evidence of a difference in vaccine effectiveness between the two sublineages of the Omicron variant.150 Indeed, the REACT-1 study of covid-19 transmission concluded that BA.2 had a daily growth rate additive advantage of 0.4 compared to BA.1.151 The risk of hospitalisation does not seem to be higher for BA.2 infection in comparison to BA.1, however.152 Emerging sublineages of the Omicron variant will be required to be monitored and reported upon for the foreseeable the future.
Although the omicron variant seems to manifest in mild disease, high infection numbers could still result in high rates of hospital admission and death in those individuals vulnerable to the virus. Omicron case numbers could be beginning to peak, however. In South Africa, a 29.7% decrease in weekly covid-19 infections were reported in the week ending 25 December 2021, compared with the previous week, and the omicron wave is said to have passed.153 Concerningly, global case numbers continue to rise rapidly154 and many countries will continue to feel the pressure exerted by the wave of omicron infections.
Variants of interest
Lambda variant C.37
The lambda variant, of the C.37 lineage, was first documented in Peru in December 2020 and was designated as a variant of interest on 14 June 2021.3 This variant contains the S protein mutations D614G, L452Q, and F490S.104 The L452Q mutation, located within the RBD, enhances binding affinity to the ACE2 receptor and increases the infectivity of the lambda variant,155 while, together L452Q and F490S, increasing the variant's resistance to vaccine elicited antibody neutralisation.155 Furthermore, F490S was identified as being a high risk mutation for enhancing abilities to escape neutralisation.155
Infectivity of the lambda variant could be higher than that of the alpha, gamma, and other D614G containing variants,156 suggesting that lambda could spread more rapidly and effectively. Additionally, compared with the primary SARS-CoV-2 virus, antibody neutralisation was found to decrease by 3.05-fold for the lambda variant, higher than that for the gamma (2.33-fold) and alpha (2.03-fold) variants.156 However, findings from a preprint study suggest that the lambda variant can be neutralised by monoclonal antibodies, and that current vaccines are protective against this variant.155
Mu variant B.1.621
The mu variant, from the B.1.621 lineage, was first documented in Columbia in January 2021 before receiving designation as a variant of interest on 30 August 2021.3 This variant contains the S protein mutations E484K, N501Y, D614G, and P681H.104 Mu also contains the S protein mutation R346K, located within the RBD,104 157 which can induce large, binding, free energy changes that disrupt the binding of antibodies to the S protein and enhance the ability of the variant to escape neutralisation.158 As discussed, the E484K, N501Y, D614G, and P681H mutations have been shown to increase transmissibility80 85 87 105 109 112 120 121 and neutralisation escape,91 106 suggesting that the mu variant is likely to be more infectious than the primary strain.
Although the lambda and mu variants have been outcompeted by the delta and now omicron variants, the development and spread of these variants of interest will need to be closely monitored and studied to appreciate their pathogenicity, transmissibility, and virulence.
Vaccines
The covid-19 pandemic prompted a rapid international search for safe and effective vaccines against the SARS-CoV-2 virus. In line with previous vaccine development, including for both SARS-CoV and MERS-CoV, the S protein was a key target for covid-19 vaccine development.159 As of 24 January 2022, 33 approved vaccines are in use in 197 countries, with 10 vaccines approved for emergency use by WHO (online supplemental table).4 ,115 133 160–251 As of 25 January 2022, 194 vaccines were in pre-clinical development and 140 were in clinical development.252 Numerous studies have explored the effectiveness of approved vaccines; however, large variations in vaccine effectiveness are reported. This variability is probably due to several factors in the studies, including the country, date, and population size of the study, as well as the SARS-CoV-2 variants circulating during the study period. These factors, along with how the effectiveness is reported, mean that it is difficult to compare vaccines and fully understand how effective each vaccine is. Here, we review the covid-19 vaccines in use around the world.
Supplemental material
BNT162b2 (Pfizer-BioNtech)
The BNT162b2 vaccine (Comirnaty) is a lipid nanoparticle formulated, nucleoside modified, mRNA vaccine encoding a modified SARS-CoV-2 S protein that was developed through a collaborative effort between Pfizer (New York, NY, USA) and BioNTech (Mainz, Germany).62 160 The vaccine was listed by WHO for emergency use on 31 December 2020253 and, as of 24 January 2022, has been approved for use in 136 countries.4
Following BNT162b2 vaccination, a response based on T helper 1 (Th1) cells is observed along with elevated levels of tumour necrosis factor α, interferon gamma, and interleukin 2, compared with placebo.254 255 The highest neutralisation titres are found between seven and 14 days after the second dose,256 while those individuals previously infected with covid-19 showed a fourfold increase in antibody binding and an 18-fold increase in neutralisation titres compared with previously uninfected individuals after two vaccine doses.257 The BNT162b vaccine is well tolerated, with limited reactogenicity. Redness and swelling at injection site have been reported, although mild or moderate pain at the injection site is the most commonly reported reaction to vaccination.256 Fatigue, muscle pain, headache, and chills are other commonly reported symptoms after BNT162b2 vaccination.258 The rate of systemic reactions after a second dose of BNT162b has been found to be 1.7 to two times higher than after a first dose, possibly suggesting an immunity boosting effect.259 Many safety reports of this vaccine describe no serious adverse events,256 259 260 but a large study of 884 828 pairs of individuals, split 1:1 based on vaccination status, found that BNT162b2 was associated with an increased risk of myocarditis, lymphadenopathy, appendicitis, and herpes zoster infection.261 Although rare, allergic reactions or anaphylaxis has also been reported after BNT162b2 vaccination.258 The online supplemental table outlines clinical trial and real world data for vaccine effectiveness.115 133 160–251
ChAdOx1 nCoV-19 (Oxford-AstraZeneca)
The ChAdOx1 nCoV-19 vaccine (AZD1222, Vaxzevria) is a non-replicating vector of the chimpanzee adenovirus ChAdOx1, modified to encode the SARS-CoV-2 S protein.262 Developed through collaboration between the University of Oxford and AstraZeneca (Cambridge, UK), this vaccine was listed by WHO for emergency use on 16 February 2021,253 and has been approved for use in 137 countries, as of 24 January 2022.4 WHO has granted emergency use listing to two versions of this vaccine (AZD1222 and Covishield) in order to use Covishield as part of their worldwide COVAX initiative, which is being produced by the Serum Institute of India and AstraZeneca-SKBio (Republic of Korea).263
Following ChAdOx1 nCoV-19 vaccination, substantial antibody production (predominantly of IgG1 and IgG3 subclasses) is seen, as well as a Th1 cell response with increased expression of interferon γ and tumour necrosis factor α.122 264 One dose of the ChAdOx1 nCoV-19 vaccine has been shown to produce a neutralising antibody response in 91% of participants, while a second dose has resulted in 100% of participants producing neutralising antibodies.265 Mild and moderate itchiness, pain, redness, swelling, tenderness, and warmth are common local reactions, while chills, fatigue, fever, headache, muscle ache, and nausea are commonly reported systemic reactions after vaccination.265 Rare symptoms, including severe chest pain, nasal bleeding, and allergic reactions have also been reported after vaccination.266 The online supplemental table outlines clinical trial and real world data for vaccine effectiveness.115 133 160–251
Ad26.COV.2.S (Johnson & Johnson)
The Ad26.COV.2.S vaccine is a non-replicating adenovirus vector, modified to contain the SARS-CoV-2 S protein in a pre-fusion stabilised conformation and requires only one dose.161 This vector was developed from the recombinant human adenovirus type 26 by the Janssen pharmaceutical company Johnson & Johnson (New Brunswick, NJ, USA),161 and was listed by WHO for emergency use on 12 March 2021.253 As of 24 January 2022, Ad26.COV.2.S has been approved for use in 106 countries.4
The Ad26.COV.2.S vaccine induces the production of a variety of antibody subclasses, such as immunoglobulins G, M, and A, and promotes several non-neutralising antibody responses, including the activation of CD4 and CD8 Th1 cells and the production of interferon γ, interleukin 2, and tumour necrosis factor α.267 268 Although neutralising antibody responses induced by the vaccine are reduced against SARS-CoV-2 variants, non-neutralising antibody and T cell responses have been found to be preserved against variants of concern,267 and a prior covid-19 infection significantly increases levels of S protein binding antibodies, antibody dependent cellular cytotoxicity, and neutralising antibodies against variants of concern (including the beta and delta variants).269 Ad26.COV.2.S is safe and well tolerated. In a large clinical trial, where 19 630 participants received Ad26.COV2.S and 19 691 received placebo, headache, fatigue, and myalgia were the most common systemic reactions, while pain at the injection site was the most common local reaction after vaccination.161 Like other vaccines, Ad26.COV.2.S has been associated with serious adverse events, such as allergic reactions and cerebral venous sinus thrombosis; however, these events are rare.258 270 The online supplemental table outlines clinical trial and real world data for vaccine effectiveness.115 133 160–251
mRNA-1273 (Moderna)
The mRNA-1273 vaccine (Spikevax) developed by Moderna (MA, USA) is a lipid-nanoparticle encapsulated mRNA vaccine expressing the SARS-CoV-2 S protein that has been pre-fusion stabilised.162 This vaccine gained WHO approval for emergency use listing on 30 April 2021,253 and as of 24 January 2022, has been approved for use in 85 countries.4
The mRNA-1273 vaccine elicits a strong CD4 Th1 cell response, with tumour necrosis factor α, interferon γ, and interleukin 2 expression increased following vaccination,271–273 while neutralising antibody titres have been shown to increase up to until around 28 days after the second vaccine dose, and remain consistently high after that.274 Fatigue, muscle pain, headache, chills, joint pain, and pain/reaction at the injection site are common adverse effects caused by the mRNA-1273 vaccine,162 258 while serious adverse effects are often avoided.162 274 Serious adverse events, including allergic reaction and anaphylaxis, are rare but not inconceivable after mRNA-1273 vaccination.258 The online supplemental table outlines clinical trial and real world data for vaccine effectiveness.115 133 160–251
Other covid-19 vaccines listed by WHO for emergency use
In addition to the covid-19 vaccines described above, five other vaccines have gained emergency use listing by WHO. Firstly, the Sinopharm BBIBP-CorV covid-19 vaccine (Covilo) was developed by the Beijing Bio-Institute of Biological Products, a subsidiary of China National Biotec Group, and was approved by WHO for emergency use on 7 May 2021.253 This vaccine is made from the SARS-CoV-2, 19nCoV-CDC-Tan-HB02 strain, which is produced in Vero cells, inactivated by β propiolactone, and then purified and absorbed with aluminium hydroxide.275
Next, the CoronaVac vaccine, developed by Sinovac Biotech (Beijing, China), was listed for WHO emergency use on 1 June 2021.253 Like the BBIBP-CorV vaccine, this vaccine is a Vero cell based, aluminium hydroxide adjuvanted, beta propiolactone inactivated vaccine, but it is based on the SARS-CoV-2 CZ02 strain.276 Covaxin (BBV152) is a whole virion inactivated, SARS-CoV-2 vaccine formula developed by Bharat Biotech International (India),277 which gained approval for emergency use listing from WHO on 3 November 2021.278
Lastly, Covovax and its originator, Nuvaxovid (NVX-CoV2372), were both developed by Novavax (MD, USA) and the Coalition for Epidemic Preparedness Innovations (Oslo, Norway), and were listed by WHO for emergency use on 17 and 21 December 2021, respectively.279 280 Both vaccines are manufactured by the same technology, and consist of a recombinant SARS-CoV-2 S protein nanoparticle combined with the adjuvant Matrix-M as a coformulation.281 These vaccines produce similar immune responses to those already discussed. Studies assessing the efficacy of these vaccines are outlined in the online supplemental table.115 133 160–251
Other approved covid-19 vaccines
In addition to the vaccines that have received emergency use listing from WHO, vaccines around the world have been developed, tested, and approved to prevent covid-19 infection. As of 24 January 2022, 33 vaccines, including those described above, have been approved in at least one country.4 The remaining 23 approved vaccines are outlined in table 2.
Summary of vaccine efficacy across vaccines approved by WHO for emergency use
Waning immunity and boosters
Throughout the covid-19 pandemic, emerging variants have threatened the effectiveness of vaccines (online supplemental table).115 133 160–251 Simultaneously, waning immunity after vaccination questions how long vaccines remain effective and highlights the importance of booster doses. Indeed, protection against SARS-CoV-2 after vaccination decreases over time, both in terms of antibody titres282–284 and vaccine effectiveness.163 285–287 However, cellular responses, such as T cell immunity, could persist for longer periods.288 289 With a gradual loss of protection from SARS-CoV-2 after covid-19 vaccination, many countries are now rolling out booster programmes with the aim of raising levels of immunity.
Since booster programmes began, evidence that a booster vaccine dose enhances antibody and cellular responses has accumulated. After a third dose of vaccine, neutralising antibody titres increase considerably290–293 and, in some cases, to higher levels than after the primary two doses.290 Additionally, boosters have also been found to increase neutralising antibody titres against the beta, gamma, delta, and omicron variants.291 294 295 T cell response is also enhanced after a third dose.292 296 297 Together, enhancing neutralising antibody and cellular responses with a booster vaccine dose is likely to provide a greater level of protection than relying on immunity built through a primary regimen.
The antibody and cellular responses observed after booster vaccinations have been found to correlate with increased levels of protection against SAR-CoV-2 infection and severe illness. On 30 July 2021, Israel was the first country to offer a third dose of BNT162b2 to certain groups. Subsequently, several observational studies have shown that those individuals who received a third vaccine dose were significantly less likely to be infected or have severe disease with SARS-CoV-2 than those who received two doses.298–301 In those individuals aged 60 or older, an observational study showed that the rate of severe covid-19 and death was lower in the group that received a booster by a factor of 17.9 and 14.7, respectively, than in the group that did not receive a booster.302 Booster doses of covid-19 vaccine have been shown to be effective against infection with the delta303 304 and, to a lesser degree, omicron variants75 145 146 304–306 despite the numerous mutations harboured by these variants. Overall, increasing evidence is pointing towards the benefits of booster doses of covid-19 vaccines; therefore, it is expected that booster programmes will continue to roll out across the globe. Based on current evidence, the US Centers for Disease Control and Prevention recommend that the time interval for receiving a booster after the primary regimen is five months for the BNT162b2 primary regimen, six months for the mRNA-1273 primary regimen, and two months for the Ad26.COV2.S primary regimen.307 As the pandemic progresses and new variants emerge, variant specific vaccines could require development, with pre-clinical studies demonstrating their efficacy308 and pharmaceutical companies, such as Pfizer, advancing in variant specific vaccine development.146 Policy makers should also consider when vaccine boosters will be given in the future and who will receive booster doses in the long term.
Emerging treatments
As the virus becomes better understood, the therapeutic strategy against covid-19 develops. Over 2000 ongoing trials are currently assessing certain treatment strategies for covid-19.309 Recently, antiviraldrugs including molnupiravir (Lagevrio) and nirmatrelvir/ritonavir (Paxlovid) have been approved in the UK,310 311 US,312 313 and Europe314 315 for treating covid-19 in certain risk groups. Similarly, sotrovimab (Xevudy), a monoclonal antibody treatment, has recently been approved for use in treating certain patients with covid-19 in the UK,316 US,317 and Europe.318 These drugs have been shown to be effective at preventing poor clinical outcomes, including death, in those individuals vulnerable to severe covid-19 infection. Other drug treatments, such as janus kinase inhibitors, corticosteroids, and anti-inflammatory drugs, have contrasting evidence to support their use; therefore, the use of specific drugs is either recommended for or against by certain treatment and management guidelines, which are discussed below.
Guidelines
The treatment and management of covid-19 is a continually evolving topic; however, health authorities have published and continue to update guidelines and recommendations for treating covid-19. The WHO living guideline on covid-19 and treatment is regularly updated, with the latest version (published on 14 January 2022) containing 14 recommendations on covid-19 treatment.319 The UK National Institute for Health and Care Excellence320 and Medicines and Healthcare products Regulatory Agency321 provide updated guidelines on covid-19 treatment, and in Europe, the ECDC regularly publishes several guidelines providing recommendations on a range of covid-19 related topics.322 The US National Institutes of Health323 and Centers for Disease Control and Prevention324 provide guidance on covid-19 treatment and management, with the Centers for Disease Control and Prevention supplying guidelines for specific groups such as employers, schools, health departments, and governments.
Considerations for the future
Novel infectious diseases and pandemics are an unpredictable but inevitable aspect of nature; therefore, we should learn from past pandemics to prepare for future ones. Firstly, the covid-19 pandemic has highlighted and amplified the existing inequalities within society,325 with Black ethnicity, social disadvantage, and unemployment being risk factors for covid-19 infection326 and those groups most economically deprived found to be particularly vulnerable.327 These inequalities need resolving in order for us to be better prepared for similar situations in the future.
Next, to progress through a pandemic we should be racing against the pathogen, and not against each other. This statement becomes apparent when considering the problems faced by countries seeking out personal protective equipment,328 and the vaccine inequity seen worldwide,329 with developed countries often better placed to be able to purchase these items. Initiatives such as WHO’s COVAX programme are vital to protect the most vulnerable groups and reduce the global spread of disease. In October 2021, the UK government released a publication outlining where the policies implemented to reduce the impact of the covid-19 pandemic failed, and the lessons learnt from these failures.330 The publication then presents conclusions and recommendations on how to enhance pandemic preparedness, lockdown and social distancing measures, testing and contact tracing, social care, and vaccines. In countries such as the UK, US, and much of Europe, where the covid-19 death rate has been high, steps need to be taken and lessons need to be learnt in order to be better prepared for the next pandemic. The responsibility of improving pandemic response lies with policy makers, the medical/scientific community, and the public, and will ultimately require a collaborative approach.
However, certain aspects of the response to the covid-19 pandemic have been a triumph. One major victory was the rapid development and rollout of vaccines,331 which continue to be effective. The rollout of rapid testing and quarantine for infected individuals was also important to at least disrupt the spread of the virus, especially given that asymptomatic individuals can contribute to the spread. Furthermore, the swift identification and sharing of knowledge of SARS-CoV-2 variants between countries should be applauded. Lessons can be learnt from countries where covid-19 was controlled. In Taiwan, authorities managing the pandemic as directed by pre-covid-19 pandemic plans prompted an immediate response. Screening of all airline passengers arriving from Wuhan and high risk areas, restricting entry for non-Taiwanese citizens, 14 day quarantine periods for contacts of people with confirmed covid-19 or returning travellers, a ban on large gatherings, and widespread mask wearing were some of the quickly implemented management strategies.332 New Zealand implemented similarly effective restrictions, with the addition of a national lockdown.332 Many of the pandemic control components that kept infection and death numbers low in Taiwan and New Zealand could be adopted by other countries in the future and could lead to improved outcomes in terms of protecting the health of individuals and the health and wellbeing of the country. Overall, much can be learnt from the covid-19 pandemic and, as we emerge from it, the inspection of which policies failed and which succeeded is imperative.
Conclusion
Covid-19 remains prevalent and life threatening. Although the rollout of vaccines has been successful, attaining a high global vaccination coverage and ensuring that all healthcare systems have the capacity to cope with seasonal waves are essential. With the omicron variant highly prevalent, we must continue to learn, develop therapeutics, and remain vigilant to new variants of concern. Here, we have provided an overview of the virology of SARS-CoV-2, including the mutations harboured by variants of the virus and how these mutations effect its transmissibility and virulence. We have also discussed the vaccines that have been developed and used around the world and have provided evidence supporting the rollout of booster doses. Future priorities should focus on continuing vaccination programmes and developing variant specific vaccines as new mutations emerge. This strategy, along with the expansion of our knowledge of SARS-CoV-2 and which treatments are most successful to treat covid-19 infections will ultimately lead to favourable outcomes.
Questions for future research
How will the SARS-CoV-2 virus mutate in the future, and which mutations will give a competitive advantage that will allow the virus to inflict disease to many people?
How do we keep up with the rapidly changing SARS-CoV-2 environment and ensure that vaccines remain effective?
How do we manage the booster programme and when will future booster vaccinations be required in order to maintain high levels of immunity?
How can we learn from the current and past pandemics so that we are better prepared for the next one?
Patient involvement
The BMJ did not request patient input on this article when it was commissioned.
Data availability statement
Data are available in a public, open access repository.
References
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
- ↵
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
MY and HC contributed equally.
Contributors MY and HC performed the literature search and drafted the manuscript. HC revised and finalised the manuscript. JS reviewed and revised the manuscript. PE was responsible for the concept and design of the work. PE reviewed, revised, and finalised the manuscript. PE is the guarantor.
Funding The authors have not declared a specific grant for this research from any funding agency in the public, commercial, or not-for-profit sectors.
Competing interests We have read and understood the BMJ policy on declaration of interests and declare the following interests: PE was funded by the UK Medical Research Council and now by Higher Education Funding Council for England, received grants from Alzheimer’s Research UK, Alzheimer’s Drug Discovery Foundation, Alzheimer’s Society UK, Medical Research Council, Alzheimer’s Association US, Van-Geest Foundation, and European Union grants; PE is a consultant to Roche, Pfizer, and Novo Nordisk; received educational and research grants from GE Healthcare, Novo Nordisk, Piramal Life Science/Life Molecular Imaging, Avid Radiopharmaceuticals and Eli Lilly; and is a member of the scientific advisory board at Novo Nordisk.
Provenance and peer review Commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.